July 8, 2026
The Microbiome Nobody Explained: Vaginal and Uterine

Your vagina isn't meant to be sterile, it's meant to be balanced. Here's what the vaginal and uterine microbiome actually do, and how fertility is affected


"Trying to conceive" sounds like it starts with a decision. It's a conversation between partners, maybe after a positive test on an ovulation predictor strip. It might be a bit spontaneous, or maybe it includes very carefully timed sex.
But the biological process that ends in pregnancy started decades before that conversation, long before that pee strip, and certainly took much longer than that all-too-brief act of coitus did. It started before you or your partner were born, inside your own developing fetuses, inside your mothers' bodies, when your early germ cells were (hopefully) already beginning to form.
Between then and now, between your own birth and the birth of your potential child, a precise sequence of biological steps has to happen in three distinct individuals (you, your partner, and your offspring), spanning fetal development, puberty, the menstrual cycle, sperm production, fertilization, and ultimately implantation — the act of officially becoming pregnant.
In this article (which is going to feel complicated but is actually oversimplified), we'll walk through 42 of those biological processes, what they're called, and what they mean, to help you understand what needs to go right, and by implication, a few of the things that can go wrong along the way.
Knowing these steps won't make any of them happen faster, and it won't make the waiting easier. But understanding the biology can reframe what "trying" actually means: You're not struggling with something simple. You're attempting to coordinate one of the most complex sequences in human biology. And most of it is invisible.
We'll do our best to explain in clear language, with everything grouped by developmental phase. But if you don't remember your highschool biology classes, this might be a bit of a tough one to follow along with.
This article is ideal for you if you're a know-it-all, an over-thinker, a details person, or trying to pinpoint exactly where your early pregnancy journey is at. But if details are going to stress you out, trust your body and save yourself 20 minutes of reading.
Because ultimately, before you see the "two pink lines" that confirm a pregnancy, there's a lot that has to go more or less perfectly, totally out of sight, down at the cellular level.
Decades before ovulation, inside both parents-to-be, who is still themselves fetuses, cells are busy preparing for reproduction in the same way they have for countless generations:
Primordial germ cell (PGC) specification and migration. PGCs are the embryonic precursors of every egg and every sperm. They arise in the yolk sac wall around 23 to 26 days post-fertilization and migrate through the embryo's hindgut to colonize the gonadal ridge (the structure that will become either an ovary or a testis).
Gonadal sex determination. Once PGCs arrive at the gonadal ridge, the embryo's sex chromosome complement (in most cases, XX for female or XY for male) drives early sexual differentiation. A cascade of proteins determines whether the gonad becomes an ovary or a testis.
♀ Oogonial proliferation. After settling in the developing fetal ovary, primordial germ cells differentiate into oogonia (developmental precursors to eggs), which replicate repeatedly through mitotic division. By the 5th month of gestation, the population peaks at around 7 million oogonia. Even after fetus is born and grows up, most of those oogonia, of course, will never get fertilized.
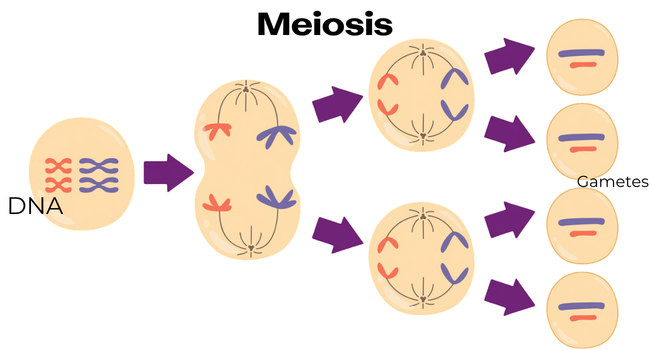
♀ Primary oocyte formation and meiosis I entry. At about 11 to 12 weeks of gestation, fetal oogonia enter meiosis, a process of DNA division, and develop into primary oocytes, thus beginning the process of halving their chromosome number in preparation for eventual fertilization.
♀ Meiotic arrest at prophase I (dictyate arrest). Primary oocytes become encased by pregranulosa cells and remain stuck at the diplotene stage of prophase I during meiosis — some for many decades — until hormonal signals in adulthood, at ovulation, trigger the process to resume. This prolonged pause is a key reason egg quality declines with age.
♀ Primordial follicle assembly. This establishes the entire lifetime supply of potential egg cells before birth, meaning a female is born with a finite, non-renewable pool. No new primordial follicles form after this point. The reserve you're born with is the total supply you'll ever have.
For women and for men, parallel preparation tracks begin at puberty, and continue for months to years before being leveraged in any future attempt at reproduction. Let's start with the guys:
♂ Spermatogonial stem cell renewal and proliferation. Male gamete development differs significantly from female gamete production in one big way: Spermatogonia in males retain the ability to divide mitotically throughout an adult male's life, beginning at puberty. These spermatogonial stem cells (SSCs) self-renew through regular mitotic divisions, continuously maintaining the stem cell pool until puberty, when they also begin producing sperm precursor cells. In other words, while "old sperm" can become an issue, men don't have an intrinsically limited sperm supply.
Moving forward from early prenatal development and puberty, fathers-to-be take a big leap forward in time. In fact, nothing important really happens at all, until approximately 60 to 74 days before conception, as spermatogenesis matures spermatogonia into sperm cells capable of carrying DNA to an egg.
Sperm meiosis (I and II). Spermatogonia undergo two sequential cell divisions. During meiosis I, sets of chromosomes pair up and exchange genetic material in a process commonly called "crossing over" (recombination), which generates genetic diversity. This is essentially the reason that siblings are still genetically different. Meiosis II then separates these chromatid pairs, yielding four haploid spermatids from each primary spermatocyte.
Spermiogenesis (spermatid maturation into spermatozoa). Next, each haploid spermatid undergoes a dramatic structural transformation. Three key changes occur: the Golgi apparatus condenses into an acrosome (a cap of enzymes needed to penetrate the egg—more on this later), the flagellum (sperm "tail") assembles for motility, and the nucleus condenses tightly as histones are replaced by protamines (basically, DNA collapses tightly in on itself for protection during transit.
Spermiation (release of mature sperm into the tubular lumen). Once spermiogenesis is complete, the mature spermatozoon detaches from its Sertoli cell (a sort of supportive nurse cell), exits the testis, and enters the seminiferous tubule. Fluid flow eventually carries it toward the epididymis, but the sperm is still not yet capable of fertilization.
Epididymal transit and maturation. This often-overlooked phase covers the time sperm spend transiting the epididymis after leaving the testis. Sperm only develop the ability to self-propel during their passage through the epididymis. At this stage, surface glycoproteins are remodeled, protective proteins are added to the plasma membrane, the cytoplasmic droplet is shed, and the sperm's ability to recognize and bind the egg's zona pellucida is established (more on this later!). Sperm are stored in the epididymis until ejaculation (see "sex", below), remaining viable for weeks in a semi-dormant state.
In all of this, it might seem like millions of perfect like sperm are being formed every day. But the truth is much messier. Each of these processes is deeply flawed and prone to failure. For each viable sperm, many more are lost to improper DNA organization, flawed sperm structures, and failed migrations. For men, fertility is much more about quantity than it is about quality. In fact, the threshold for "normal" sperm motility is just 40% — meaning the majority of sperm in any given sample aren't expected to be viable.
Combined, the processes of spermatogenesis (sperm production and epididymal maturation) are quite lengthy: it takes about 74 days to transform spermatogonial cells into sperm and transport them to the epididymis, with another 12 days needed for epididymal maturation. Which means that today's good or bad habits won't be fully reflected in semen analysis (or conception attempts) until about three months from now.
And for that conception attempt? Assuming you've got the birds and the bees more or less figured out, or have a method of science-ing your way around that part, you're still going to need an all-critical female partner. Which brings us to:
This group covers the approximately 10 to 14 days before ovulation in a post-pubescent woman, during which the body (usually) selects a single dominant follicle.
Endometrial Proliferation. From menstruation through to ovulation, rising estrogen from growing follicles drives the endometrium to proliferate. Glands lengthen, stromal cells divide, and the functional layer thickens from approximately 1 mm to 7–14 mm, creating the structural foundation for later fetal implantation.
Follicle activation. Way back up at the top of this article, in the section of the developmental origins of pregnancy in the parental fetus, we ended with "Primordial follicle assembly." To recap: At birth, we've got this huge pool of potential eggs. Then, beginning at puberty, small numbers of primordial follicles are continuously "activated" from the dormant pool by local intra-ovarian signals (including one called the PI3K-PTEN-mTOR pathway).
Early follicle development. Once activated, follicles progress over many months through primary, secondary, and early antral stages, growing and acquiring the capacity to respond to gonadotropin hormones. Again, this starts happening many months before the hormone-driven ovulatory cycle they'll eventually be a part of.
Follicle recruitment and intercycle selection. Finally, at the start of a new menstrual cycle, rising follicle-stimulating hormone (FSH) from the pituitary gland stimulates a cohort of developed antral follicles to begin to grow. As they grow, they race to secrete both estrogen and inhibin B, which prevent the growth of follicles around them. In this way, the follicles with the most FSH receptors grow the fastest, and best inhibit the growth of their follicle peers, until (usually) only one remains. This winner — the dominant follicle — is typically 18 to 25 mm in diameter (about the size of a marble!) and is the one designated to ovulate.
Estrogen rise and LH surge. As the dominant follicle grows, it also secretes estradiol. Once estradiol levels remain elevated for approximately 36 to 48 hours, the pituitary gland responds with a dramatic spike of luteinizing hormone (LH). This is the famous LH surge that ovulation predictor kits (OPKs) try to detect. This surge will trigger the remaining ovulation steps:
These steps unfold over approximately 36 to 40 hours following the LH surge.
Oocyte meiosis resumption. That dominant follicle, finally allowed to grow beyond its primordial follicle assembly, contains a single oocyte. But the oocyte itself has been stuck at meiotic prophase I (the dictyate arrest) since before the mother was born. The LH surge finally breaks this dictyate arrest. The oocyte's nuclear envelope (the "germinal vesicle") dissolves, chromosomes condense and separate, and meiosis I completes. The oocyte enters meiosis II but then pauses again(!) at metaphase II. It will only finish the process if fertilization occurs.
Cumulus expansion and follicle wall remodeling. A cloud of granulosa cells (the "cumulus oophorus") surround the oocyte. The granulosa cells secrete a "hyaluronic acid-rich extracellular matrix"—basically a bunch of goop—which causes the cumulus complex to expand dramatically. At the same time, proteases and prostaglandins degrade the follicle wall at the "stigma" — the site where the egg will soon burst out of the ovary.
Not clear? Imagine the egg is a marble, and it's inside the follicle, which is a balloon. The balloon has been growing because it's getting filled with water. But at the same time, someone is using sandpaper to scratch the knot that tied the balloon shut. So the balloon/follicle is stretching, and there's a weak point. It's destined to burst, and it will eject the marble/egg.
Follicle rupture and COC expulsion. Approximately 36 to 40 hours after the LH surge, the follicle wall ruptures. The cumulus-oocyte complex (COC) — the egg surrounded by its cloud of cumulus cells — is expelled into the peritoneal cavity near the fallopian tube opening. This is the moment of peak fertility.
So now we've got an egg hanging out near a fallopian tube. But a lot needs to happen in a pretty short span of time in order to have any chance of making a baby:
COC capture and transport. The fimbriae (finger-like projections at the fallopian tube opening) catch the COC (the marble) and sweep it into the tube. Ciliary beating and muscular contractions carry it toward the ampulla — the fertilization site — over a period of a few hours. In total, it will travel 10–12cm (4 to 5 inches).
Corpus luteum formation and progesterone secretion. The collapsed follicle (the burst balloon) transforms into the corpus luteum, a temporary endocrine structure that secretes progesterone. Progesterone shifts the endometrium (the lining of the uterus) from the "proliferative" phase to the "secretory" phase, preparing it for implantation. In about 14 days, if there's no embryo, the corpus luteum will degenerate, progesterone will fall, and menstruation will occur. But if there's an embryo, the corpus luteum will stick around for most of the first trimester, helping to sustain the pregnancy with progesterone, until the placenta is fully activated.
Endometrial secretory phase transformation. After ovulation, progesterone from the corpus luteum transforms the endometrium. Uterine glands begin secreting histotroph (nutritive fluid). And on the uterine surface, specialized epithelial projections called pinopodes form. These finger-like structures are thought to aid blastocyst adhesion, and serve as a specific marker of endometrial readiness for pregnancy.
The part you think about the most, because it's in your direct control, but it's also low-key the least complicated, at least physically speaking.
♂ Ejaculation and semen liquefaction. During ejaculation, sperm from the epididymis mix with seminal vesicle fluid, secretions from the prostate, and urethral gland secretions to form the mixture we call semen. Semen is initially quite thick (due to seminal vesicle proteins), but prostate-derived proteases (PSA) will cause it to liquefy, so that within 5 to 30 minutes, sperm can swim freely.
Sperm transport. Assuming you had barrier-free penis-in-vagina sex, of the hundreds of millions of sperm ejaculated, still only a few hundred will successfully make it through the cervical mucus and enter the uterus. This is a key way that IUI is useful: Sperm transport is partially skipped. In IUI, the doctor will use a catheter to bypass the cervix and put more sperm directly into the uterus. In both natural conception and IUI, subtle uterine contractions help propel sperm toward the fallopian tubes, where they will attach to epithelial cells at the isthmus, forming a sort of sperm reservoir. From there, sperm are released deeper into the fallopian tube in waves, to attempt a meet up with the oocyte.
Sperm capacitation. Fun fact: Sperm, as ejaculated, literally cannot fertilize an egg. Capacitation is a molecular transformation that occurs as a result of uterine secretions, and makes it all possible: In short, the sperm's cell wall is weakened, and its flagellum's movements are accelerated. Only in this state of "hyperactivated motility" is the sperm able to move through viscous environments and transfer its DNA.
Sperm chemotaxis and thermotaxis. Capacitated sperm navigate toward the egg using two directional signals: (1) a temperature gradient of approximately 2°C between the cooler isthmic reservoir and the warmer ampullary fertilization site (called thermotaxis), and (2) progesterone and other chemical attractants secreted by the COC that activate ion channels and help sperm steer (chemotaxis).
These steps occur in the approximately 12 to 24 hours after ovulation, inside the fallopian tube, when a wave of sperm encounters the COC (the cumulus-oocyte complex; that egg wrapped in a cloud of cells).
Zona pellucida penetration. First, the sperm use an enzyme to disperse the cumulus matrix. Then, they attempt to use the ZP3 glycoprotein to bind to the egg in an area called the zona pellucida ("transparent zone"). If successful, this binding triggers the acrosome reaction, a release of enzymes from the sperm's acrosomal cap (remember? it formed during spermiogenesis just for this reason!), which, combined with the hyperactivated motility, enables the sperm to physically drill through the zona.
Gamete membrane fusion (IZUMO1–JUNO). If the sperm makes it through the zona, it reaches the oocyte plasma membrane (called "oolemma"). At this point, the sperm surface protein IZUMO1 binds to JUNO, the corresponding receptor on the egg's surface. This is the first true physical link between the two cells, and is necessary for full membrane fusion. Both proteins are essential for fertility, and apparently nothing else: In tests on mice, eliminating the gendered protein resulted in a otherwise healthy—but complete infertile—mouse.
Polyspermy block. Within seconds of gamete membrane fusion, the oocyte releases cortical granules from beneath its membrane. These granules release enzymes (including ovastacin) that modify and harden the zona pellucida, blocking additional sperm from penetrating. Simultaneously, JUNO are rapidly shed from the oolemma, removing the adhesion receptor that allows sperm to fuse to the egg, creating a secondary membrane block.
Pronucleus formation. Sperm entry triggers calcium oscillations in the oocyte that activate meiosis II resumption and completion (finally!), forming the female pronucleus. Meanwhile, the tightly-wound-for-transport sperm nucleus decondenses, forming the male pronucleus.
Syngamy. The male and female pronuclei migrate toward each other, their envelopes break down, and the chromosomes — 23 from each, for 46 in total — condense onto the first mitotic spindle. We now have a diploid genome: Two cells become one! The first cellular division of the new zygote follows immediately. This is fertilization! But there's a long ways to go from here.
This group covers days 1 to 4 after fertilization, as the embryo travels down the fallopian tube while dividing.
Early cleavage — first divisions, embryonic genome activation, and compaction. The zygote undergoes rapid mitotic divisions (cleavage) in which cells (blastomeres) halve in size without the embryo growing. Around the 4-to-8-cell stage, the embryonic genome activates for the first time (embryonic genome activation, or EGA), switching control from maternally stored RNA to the embryo's own gene expression. At the 8-to-16-cell stage, blastomeres flatten against one another and tight junctions form — a process called compaction.
Morula formation. By day 3 to 4, the compacted embryo is a solid ball of approximately 16 to 32 cells called a morula. It's still enclosed within the zona pellucida and has not yet implanted in the uterus. Fluid begins to accumulate between cells, setting the stage for blastocyst cavitation.
Days 4 to 6 after fertilization bring the early embryo's most dramatic structural reorganization.
Blastocyst formation — cavitation, ICM and trophectoderm specification. Fluid pumped into the morula coalesces into a central cavity called the blastocoel, transforming the morula into a blastocyst. As this happens, cells sort into two distinct lineages: the inner cell mass (ICM), which will become the embryo proper and yolk sac, and the trophectoderm, which will become the placenta. This is the first major cell specialization of human development.
Zona hatching and uterine entry. The expanding blastocyst exerts mechanical pressure on the zona pellucida, which thins and ruptures around day 5 to 6. The blastocyst "hatches" free of its egg, and enters the uterine cavity. Its trophectoderm surface is now exposed and able to interact directly with the endometrium.
This final group covers days 6 to 10 after ovulation and takes the process from the hatched blastocyst's arrival through to the establishment of early placental circulation.
Implantation window opening. The limited period of optimal endometrial receptivity is commonly referred to as the "window of implantation" and is generally detected between days 20 and 24 of a normal 28-day menstrual cycle (typically between the 6th and 10th day after ovulation). This is a relatively short period where the endometrial lining is most conducive to allowing an embryo to implant. Outside of this window, implantation cannot occur. The timing of blastocyst arrival and endometrial readiness have to be synchronized. Missing the window is a recognized cause of implantation failure and recurrent pregnancy loss.
Blastocyst apposition and adhesion. The hatched blastocyst first loosely rolls along the endometrial surface (apposition) before forming stable adhesion contacts between trophectoderm surface molecules (including L-selectin and integrins) and corresponding endometrial structures. The embryo orients with the ICM pole facing the endometrium.
Trophoblast differentiation into cytotrophoblast and syncytiotrophoblast. The trophectoderm differentiates into two cell types: cytotrophoblasts (the proliferative, mononuclear stem cell layer) and the syncytiotrophoblast (a multinucleated mass formed by cytotrophoblast fusion) that actively invades the uterine lining. The syncytiotrophoblast has no cell boundaries, enabling deep tissue invasion.
Syncytiotrophoblast invasion and decidualization. The syncytiotrophoblast erodes into the endometrial stroma. In response, stromal cells undergo decidualization, which is a transformation involving enlargement, cytoplasmic glycogen accumulation, and expression of prolactin and IGFBP-1. This creates an immune-tolerant environment for the embryo, so that mama's body doesn't reject it as a foreign invader.
Interstitial implantation (blastocyst embedding). In humans, implantation is interstitial: the entire blastocyst embeds within the endometrial tissue, becoming completely surrounded by it. The endometrial epithelium actually closes over the implantation site, concealing the embryo. This process completes around day 10 after ovulation.
hCG secretion and corpus luteum rescue. Once the syncytiotrophoblast contacts maternal blood in the endometrial tissue, it begins to secrete human chorionic gonadotropin (hCG). hCG binds to the LH receptor on the corpus luteum and prevents its programmed regression. So where normally your progesterone drops at this phase of your menstrual cycle, with an implanted embryo, your corpus luteum keeps producing progesterone. While hCG is the hormone detected by home pregnancy tests, the true purpose of this molecular signal it to "rescue" the corpus luteum and sustain the pregnancy.
When hCG is detected at an abnormally high level in your blood or urine, you're officially pregnant! The "last step," other than growing and delivering a healthy baby (no small feat!), is to build a placenta.
Placentation initiation. Cytotrophoblasts continue proliferating and invading deeper into the endometrium, eventually reaching and remodeling maternal arteries. These arteries transform from high-resistance, narrow-bore vessels into wide, low-resistance channels that flood the placental tissue with maternal blood. While the placenta continues to grow and develop until the end of the first trimester and beyond, this arterial remodeling is the foundation of its functioning.
Pregnancy is an endpoint that sits on top of a huge number of biological processes, most of which have a name and an entire body of scientific research behind them. Some of these started forming before you were ever born, many of them take place over months or even decades, and some of them have to happen perfectly within a carefully timed window of only a few hours.
If you're in the middle of this — tracking cycles, timing everything, wondering why it hasn't worked yet, or why it keeps going badly — the sheer number of steps might feel overwhelming. But hopefully knowing more about what's happening also gives you a sense of validation. Because none of this is easy.
The more you understand about what's happening beneath the surface, the better positioned you are to ask the right questions, push for the right tests, and be a good advocate for yourself and for your partner.
Meiotic arrest means your primary oocytes hit pause during the first meiotic division (called dictyate, at the prophase I stage), and stay paused for decades, from before birth until the LH surge triggers ovulation. Primary oocytes remain arrested in the diplotene stage of prophase I during meiosis until the onset of puberty and beyond. The longer the arrest lasts, the greater the risk of chromosomal errors (such as non-disjunction) during meiotic resumption, which is why egg quality and the rate of aneuploidy increase significantly with maternal age.
Sperm need time in the female tract before they can fertilize an egg. Transport through the cervix and uterus takes minutes to hours, but capacitation (the molecular priming step) requires several additional hours. Once ovulation occurs, the egg remains viable for approximately one day. In practice, fertilization can occur anywhere from a few hours to about a day after ovulation, depending on whether sperm were already waiting in the isthmic reservoir, or arrived after the egg was released.
Cytotrophoblasts are the proliferative, mononuclear stem cell layer. They divide and replenish the trophoblast population. The syncytiotrophoblast is a multinucleated mass formed by cytotrophoblast fusion, with no internal cell boundaries, that actively invades the uterine lining and produces hCG. This distinction matters because the syncytiotrophoblast is the tissue in direct contact with maternal blood and is responsible for both hormone production and nutrient exchange, which is the functional foundation of the placenta.
In healthy adult males, the whole spermatogenesis process takes between 64 to 74 days. Once formed, sperm take another 8 to 12 days to pass through the epididymis, where they mature and develop motility. That means the sperm in today's ejaculate started developing roughly 3 months ago. So a dietary change, supplement, sauna habit, or illness today won't show up in a semen analysis for about 3 months. That's also the timeline fertility specialists use when evaluating whether interventions are working.
Endometrial receptivity refers to a short period when the endometrial lining is most conducive to allowing an embryo to implant, typically between the 6th and 10th day after ovulation. Missing this window or having a non-receptive endometrium during this period can result in failed implantation, even if a high-quality embryo is present. Several studies have shown that the window of implantation may differ among women, and a personalized window that is displaced has been identified in about one out of four patients suffering from recurrent implantation failure. This is why synchrony between embryo development and endometrial readiness is essential for pregnancy success.
Download myStoria today (it's free) and record a voice note talking through your recent reproductive health history. The app will automatically create a timeline of your health information, which provides critical context that Luna (our reproductive health chat assistant) uses to give you personalized answers to all of your questions.
Need more help after that? Use the app to contact a live human health concierge.

Paul Baribeau is a growth and lifecycle marketer at myStoria. He writes about health and benefits from an unlikely vantage point: a grounding in pure science, psychology, and philosophy of science; hands-on experience in residential care and HR technology; and years navigating the system as a patient and advocate.